Several phenomena are expressed differently
or are of another order of magnitude in an acceleration field (e.g. 1×g
Earth's gravity) as compared to a near weightlessness conditions. When thinking
about any effects of gravity on e.g. a biological cell one should distinguish
between direct and indirect effects.
Direct effects may be described as effects of acceleration directly onto the
cell or its internal components. Indirect effects are effects on the cell surroundings.
Some effects reported in cell cultures experiments might be interpreted as 'direct'
while 'indirect' effects might not be excluded.
When considering gravitational research we have to be aware of several phenomena that play a role, especially at the level of a biological cell in vitro (i.e. in a 'perti-dish').
Sedimentation is the downwards movement (along the gravity vector) of an object in such a way that the object moves relative to its surrounding medium. During this process the medium, such as fluid, exerts a force on the object known as drag force. This drag force is due to the viscosity, and also, at high speeds, to turbulence behind the object.
By definition, sedimentation force (FS) is the downward force of weight due to linear acceleration (mg) minus buoyancy (FB) minus frictional forces opposing downward motion (FF).
FS = mg - FB - FV
Where:
m = mass of the object (kg)
g = the gravity acceleration (m/s2)
FB = buoyant force (see later)
= rfVg
rf = specific density of the fluid (g/cm3)
V = volume of the displaced fluid (= volume of the object) (m3)
FF = frictional force
= kv
k = the Stokes equation: 6prh (for a spherical object)
r = radius of the object (m)
h = viscosity constant (N·s/m2 or, Pa·s)
v = object's velocity relative to the fluid (m/s)
FS = mg - (rfVg) - (6prhv)
Sedimentation is one of the main differences between an on Earth and an in space experimental environment. As from every days experience, we know that big (>r) and heavy (>r) particles are the quickest settlers. These relatively big and heavy particles play a part in the positive gravitropism of plant roots. The so called amyloplasts, localized in specialized cells in the root tip, the statocytes, sediment to the bottom of the cell in response to gravity. Also, for the fresh water unicellular ciliate Loxodes it is argued that the heavy cell compartments (Mueller organelles), containing bariumsulphate crystals, may be a good candidate for gravisensor in these species.
Hydrostatic pressure is also influenced by gravity. In situations of hypergravity part of the pressure which is acting on a surface area derives from the weight of the liquid column standing above these surfaces, like culture medium above a monolayer of cells. Pressure is defined as force per unit area, where the force, F, is always acting perpendicular to that surface. The IS unit for pressure is the Pascal (Pa), one Pascal being one Newton per square meter (N/m2).
In liquids or gasses of constant density:
P = F/A
= rAgh/A
Where:
P = pressure (Pa)
F = force (N)
A = surface area (m2)
r = density of the liquid or gas (g/cm3)
g = unit gravity (m/s2)
h = height of the liquid (or gas) column (m)
Normally, the pressure exerted by a fluid column is additional to atmospheric pressure. The total pressure would be:
Ptot = rgh + Po
Where:
Po = atmospheric pressure (Pa)
It is clear from this equation that hydrostatic pressure is linearly proportional to g. Under pure weightlessness conditions, hydrostatic pressure due to weight is zero. One should keep in mind though, that in manned missions the spacecraft interior pressure exerts a hydrostatic pressure in a liquid. During hypergravity experiments, however, in which cells were cultured at 320´g,(Inoue et al. 1993) with an assumed liquid column above the cells of 1 mm, the hydrostatic pressure on the surface of the culture plate due to liquid weight would be nearly 3.3 kPa (see calculations), and almost 10 kPa with a 3 mm liquid column. In addition to the pressure deriving from the liquid column, also the atmospheric pressure should be taken into account. Atmospheric pressure, normally referred to as one atmosphere, is about 100 kPa at sea level.
P = rgh
= 1.05×103 kg/m3 × 9.81 m/s2 × 320 × 0.001 m
= 3.296 kPaWhere:
r = specific density of culture medium @ 1.05 g/cm3
g = 320 × unit gravity (m/s2)
h = 1 mm liquid column above the cell layer (m)
In the Inoue experiment the force due to hydrostatic pressure acting upon a particular cell surface in a monolayer culture with a cell diameter of 10 µm would be around 2.6·10-7 N. In comparison, the force of the same cell due to its own weight would be around 5.1·10-12 N. This indicates that hydrostatic pressures cannot be neglected when considering cells subjected to hypergravity or microgravity conditions.
According to the kinetic theories, the molecules in a gas or in a liquid, are continually moving about and are colliding with each other. This is the process of diffusion. It can also be visualized microscopically, and is better known as the Brownian movement of small particles in solution. The diffusion process is similar to the conduction of heat through a mass. The conduction of heat takes place in response to a temperature gradient while diffusion takes place in response to a concentration gradient. The physiologist Adolf Fick formulated his equation for diffusion in 1855.
DN / Dt = -DA × (DC /Dx)
Where:
DN = number of diffusing molecules (n)
Dt = time needed for diffusion (s)
A = surface area on which diffusion takes place (m2)
DC = (C1-C2) concentration difference at 2 locations within the surface area
Dx = distance between C1 and C2 (m)
D = the diffusion coefficient for a particular substance= (kT/4pha)
Where:
k = Bolzmann constant (JK-1)
T = temperature (K)
r = radius of diffusing molecule (assumed spherical) (m)
= for a spherical particle r = ((3M/(4prNa))1/3Where:
M = mass per mole
r = density of the molecule (kg/l)
NA = Avogadro's number (mol-1) (Todd 1989)
h = viscosity constant (Ns/m2 = Pa•s)
The minus sign in -DA reminds us that the flow of molecules is opposite to the concentration gradient. Diffusion is not altered under microgravity. Therefore, in the absence of sedimentation, diffusion is on the molecular level one of the main remaining in vitro 'mixing facilities' under weightlessness conditions.
Convection is the movement of fluid or gas, based on density variations. When a part of a liquid or gas is heated up it expands, causing it’s density to decrease. In the same liquid this heated part shall move up since it is lighter. This phenomenon is only present under conditions of acceleration (i.e. gravity) and is absent in weightlessness. The impact of density differences within a biological cell due to temperature differences is negligible, based on the high heat conductivity and relatively small temperature differences in cells.(Pollard 1965 The absence of convection during in vitro experiments in space could be that spent culture medium and metabolic waste products, but also autocrine growth factors, are accumulating around the cells and tissues under microgravity. This could of course modulate growth and differentiation. The impact would be a secondary effect of microgravity on cell metabolism.
The effects of diffusion and convection differ considerably in magnitudes. The following is to give an idea of diffusion velocity times for non-colliding particles. It is based on Einstein's mean-square or "random-walk", x2, diffusion distance. The diffusion constant of K+ ions in water is, D=0.001 mm2/s.(Ling 1984) For a one dimensional displacement over 5 mm by diffusion, it would take K+ ions nearly;
t = x2/D
t = 25/0.001 = 25000 s = 6.9 hrs
Where:
x = displacement distance (mm)
D = diffusion constant (mm2/s)
Some examples of diffusion rates:
Reference: * = Handbook Chemistry & Physics, ** = Todd 1989.
Particle / molecule diameter Diffusion rate (D) in cm2/sec any (spherical) 1 mm 25°C water 5×10-12* any (spherical) 1 µm 25°C water 5×10-9* any (spherical) 1 nm 25°C water 5×10-6* raffinose - 15°C water 3.3×10-6* sucrose - 25°C water 5.2×10-6* 1-butanol - 25°C water 5.6×10-6* glucose - 25°C water 6.7×10-6* ethanol - 25°C water 12.4×10-6* methanol - 15°C water 12.8×10-6* methane - 25°C water 14.9×10-6* albumine - - 6×10-7** whole cell ~10 µm - < 10-10**
Convection currents that result from density differences generate greater mixing capacity. Assume a spherical particle with a radius r = 0.1 mm in water. The density of water r = 1 g/cm3 and the viscosity h = 0.01 poise. Assume the specific gravity of the particle is 1% more than water, r = 1.01 g/cm3. The Stokes equation for friction, F, would be;
F = 6prhv = 4/3pr3(rwater-r)g
v = 2/9h(rwater-r) r2g
v = 0.2 mm/s
Where:
r = radius of particle (m)
h = viscosity constant (Pa·s)
v = velocity (m/s)
r = density of particle (g/cm3)
Or, within 6.9 hrs this particle would have traveled nearly 5 meters. In this example the convection process is nearly 1000 times faster compared to diffusion.
The phenomenon of buoyancy driven convection, or Rayleigh convection, is lacking in a weightlessness environment. Density differences that cause this convection are the result of local changes in temperature or composition. This is one of the important differences between ground (1×g) and flight (µg) experiments. The net force acting on an object in relation to it's buoyancy is:
FB, net = mg + F1 - F2
mg = downwards force due to the object's weight
F1 = downwards force due to the fluid on top of the object
= rfgh1AF2 = upwards force due to buoyancy
= rfgh2AWhere:
m = mass of the object (kg)
rf = density of the fluid (g/cm3)
g = the acceleration (m/s2)
h1 = height of the fluid column above the object (m)
h2 = height of the fluid column from the surface of the fluid to the bottom of the object (m)
A = surface area (of a cylinder or cube) (m2)
For a hypothetical cylindrical shaped object; (h2-h1)A = V, is the volume of this cylinder and rf×V= M is the mass of the fluid with a volume equal to the volume of the object.
rfg(h2-h1)A = Mg = wf
Where wf is the weight of the fluid which has been displaced by the object. So;
FB, net = mg - wf
The force of buoyancy of an object works opposite to gravity, and is proportional to the objects' volume and the liquid's weight. Buoyancy is not present under weightlessness conditions.
For a hypothetical mammalian cell with a diameter of 10 µm and a density r of 1.05 g/cm3 in a liquid culture medium of r=1.01, the upwards of buoyant force would be approximately
F = 5.24·10-10 × 1.05 × 9.81 - 5.24·10-10 × 1.01
= 4.87·10-9 N
Another phenomema ralated to buoyancy but acually completely different is Marangoni convection. It is the effect on mass transfer across a liquid-gas interface. The phenomenon of liquid flowing along an interface from places with low surface tension to places with a higher surface tension is named after the Italian physicist Carlo Giuseppe Matteo Marangoni (1840-1925). The Marangoni number is frequently expressed as:
Ma =((-dg/dc)c0H)/mD
In this equation (dg/dc) is the dependence of surface tension on concentration, c0 a characteristic concentration, H a characteristic length and D the diffusivity of the solute in the phase of interest. This Marangoni number expresses a ratio of a characteristic diffusion time and a characteristic time for Marangoni driven flow. Usually, the above mentioned buoyancy or Rayleigh effect dominates the flow in liquid layers with dimensions larger than one centimetre. The Marangoni effect usually dominates when the characteristic dimension is smaller than one millimetre. For chemical engineers, the layers of 1 mm are more relevant as these layers are more often encountered in mass transfer equipment. However, it is practically impossible to study the flow pattern in such thin layers, not even with the help of a sophisticated Laser-Doppler anemometer. Therefore, the only way to study Marangoni flows separately from buoyancy in 1 cm layers, is by experiments in a reduced gravity environment. So Marangoni convection does not change under microgravity conditions but the near weightlessness environments facilities a better measurement of this phenomenon. For elaborate informetion on the Marangoni effect you may read the academic thesis from Tjaart Molenkamp of the University of Groningen entitled: 'Marangoni convection, mass transfer and microgravity.' (See Publications / Academic Thesis).
The Coriolis acceleration is a type of g-field that acts upon moving objects within a rotating system such as the Earth or a centrifuge. Coriolis accelerations are so called cross-coupled responses, due to the angular motion in two planes. It is an effect of rotation that contributes to the impurity of the gravity generated by a rotating system. For gravitational research it is particularly involved in studies of moving objects, like flagellates in hypergravity fields. In humans, this acceleration is responsible for motion and space sickness, by action on the inner ears' semicircular canals. The definition of this acceleration in g's is:
aCoriolis = (2v × v)/g
Where:
aCoriolis = Coriolis acceleration expressed in g's
v = velocity of the moving object (m/s)
v = angular velocity of rotating system (rad/s)
g = acceleration due to gravity (m/s2)
If the angular velocity is expressed in revolutions per minute (rpm), the equation above changes to:
aCoriolis = ((2pv)/30×g) × rpm
Where:
rpm = revolutions per minute (n), centrifuge speed.
The impact of the Coriolis force upon a sample depends on the axes along which the object moves in respect to the rotation axes. The highest impact is when the motion of the sample is in a plane 90° to the rotation axes.
The impact of this phenomenon is increased in a rapidly changing angular acceleration field such as centrifuges with relatively small diameters for generating a certain g-value compared to the velocity, v, of the object studied. Also the magnitude, rpm, of the primary acceleration is important. Two rotating systems with different radii but spinning at similar rpm's would generate the same Coriolis accelerations.
For example, for an object like the unicellular Loxodes moving at a speed of 200 µm/s(Hemmersbach 1994, personal communication ) in a rotating system of 100 rpm, the Coriolis acceleration would be 4.2·10-4g. If the rotation would generate a g-force of 1.0×g, the perturbation due to the Coriolis acceleration would be less than 0.05%.
The Coriolis force is also present in the fast rotating clinostat, an instrument to mimic some aspects of real microgravity. At a clinostat speed of 60 rpm and a tube radius of 1 mm, the centrifugal force at the rim is 4.2·10-3g while the corresponding Coriolis acceleration is no more than 2·10-23g.(Briegleb 1992)
After this extended
list of possible effects that might play a role in gravitational
research as 'secondary effects' it is of course more interesting
to speculate what the direct effects of gravity on a cell might
be. To start, some of the effects listed as 'secondary' might
well also take place inside the cell. In that case we regard them
as 'primary'.
Another very interesting idea of how gravity might have an impact on small biological
systems was brought forward by Dick Mesland some 12 years ago (Mesland 1987).
Based on the publication from Prigogine and Stenger where they discuss systems
that are in 'far-from-equilibrium 'steady-states' he reasons that gravity, which
is actually a very weak force at the level of the cell, might indeed have an
impact on the cell.
In, thermodynamically, far-from-equilibrium steady-state systems only very small
perturbations within such a system might have very significant effects. They
claim there hypothesis can be seen in e.g. enzymatic reaction chains where reaction
products have direct effects on the reaction itself. Some of these systems arrive
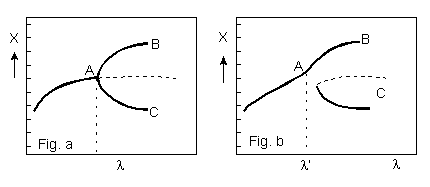
at a bifurcation point (see Figure). At this point the system has to 'choose'
between two pathways. This choice might be purely accidental, but at this point
the system becomes extremely sensitive to external stresses. Prigogine and Strenger
claim that relatively weak forces, like gravity, may have a decisive role in
such systems.
Figure: Bifurcation in a far-from-equilibrium nonlinear steady-state system. X can be the concentration of a particular reactant in the system, while l is a system parameter. In Fig. a, increasing l to l' drives the system to A where it can enter either pathways B or C with equal probability. In A the system becomes extremely sensitive to external stresses such as gravity. In Fig. b, increasing l, in the presence of an external stress results in only one possible pathway, B. (See Kondepudi & Prigogine 1983 and Prigogine & Strenger 1984)
Some references
• Albrecht-Buehler G. Possible mechanisms of indirect sensing by cells.
ASGSB Bulletin 4(2), 25-34, July 1991.
• Albrecht-Buehler G. The simulation of microgravity conditions on the
ground. ASGSB Bulletin 5(2), 3-10, October 1992.
• Briegleb W. Some quantitative aspects of the fast-rotating clinostat
as a research tool. ASGSB Bulletin Vol.5(2), Chap. 3, 23-30, 1992.
• Fundamentals of Aerospace Medicine. Edt. DeHart R.L., Lea & Febiger,
Philadelphia, 346-347, 1985.
• Giancoli D.C. General physics. Edt. L. Mihatov. Prentice-Hall Inc., New
Jersey, U.S.A. 1984.
• Inoue H., Nakamura O., Duan Y., Hiraki Y., Sakuda M. The effect of centifugal
force on the growth of mouse osteoblastic MC3T3-E1 cells in vitro. J. Dent Res.
72(9), 1351-1355, 1993.
• Ling G. In search of the physical basis of life. New York, Plenum Press,
p.115, 1984.
• Marion J.B. General physics with bioscience assays. John Wiley &
Sons Inc., New York, 1979.
• Meslamd D.A.M. Biology in Space. In: Microgravity Research. Status and
Prospects. Proceedings of the SRON Symposium 2-3 April 1987. Edt. P. van Nes
& B. Battrick. SRON, Utrecht, The Netherlands, 1987.
• Minster O., Innocenti L., Mesland D. Looking at science on board Eureca.
ESA BR-80, 3-6, May 1993.
• Otto G.H., Schatz A. Methods to obtain and to simulate low-gravity conditions
Part I. Deutsche Forschungs- und Versuchsanstalt für Luft- und Raumfahrt
(DFVLR), Köln-Porz.
• Pollard E.C. Theoretical studies on living systems in the absence of
mechanical stress. J. Theoret. Biol. 8, 113-123, 1965.
• Kondepudi D.K., Prigogine
I. Sensitivity on nonequilibrium chemical systems to gravitational field. Adv.
Space Res. 3(5), 171-176, 1983.
• Prigogine I. & Stenger I. Order out of Chaos, Man's new dialogue
with nature. Bantam Books, Toronto, New York, London and Sydney, 1984.
• Stephenson R.J. Mechanics and properties of matter. 3rd Edition. John
Wiley & Sons, Inc., 14-17.
• Todd P. Gravity-dependent phenomena at the scale of the single cell.
ASGSB Bulletin 2, 95-113, 1989.