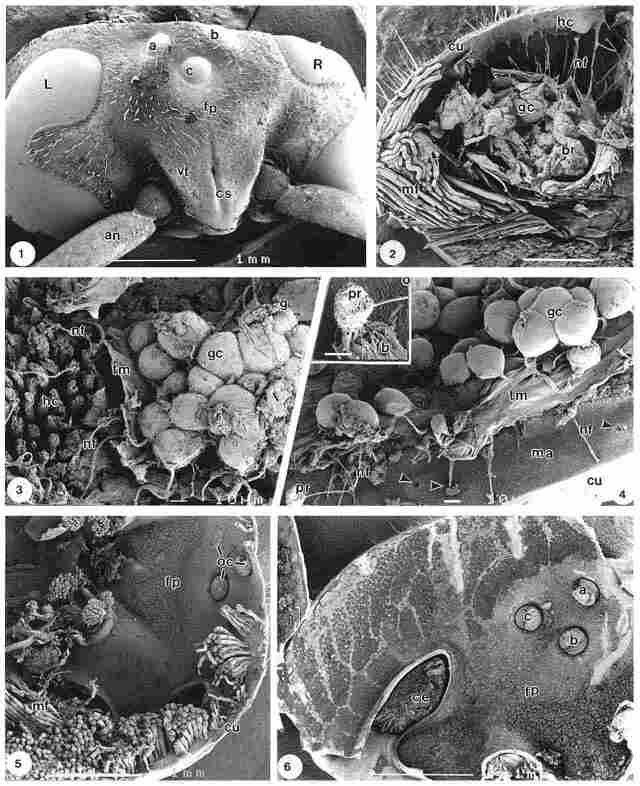
Plate I (for details see text)
CHAPTER 4
Ciliary hair cells and cuticular photoreceptor of the hornet Vespa orientalis as components of a gravity detecting system: a SEM/ TEM investigation
Abstract
This chapter describes three types of hair cell configurations with stereo-and kinocilia in the head of the hornet; these were encountered at the vertex and frons regions adjacent to the three ocelli and are assumed to be part of the hornet's gravity detecting system, together with cuticular photoreceptors. The first and most common type of hair cell configuration (type A) was a cell surrounded by a septum, having a diameter of 30-50 µm. Aggregates of over 20 of such hair cell groups together formed a larger unit, 130-300 µm in diameter, which was also enclosed by a septum. Many of these larger round units were, in turn, arranged in either angular or leaflike clusters. The hair cells bore numerous cilia of 4.5-6.0 µm long, and were themselves composed of smaller subunits of about 7-8 µm in diameter, which were not enclosed by a septum. The second type of hair cell configuration (type B) was discrete cells with a diameter of 12.5-14 µm. Type B cells are located in the vicinity of the pore canal outlet of the peripheral photoreceptor. These single hair cells were either devoid of or only partially enclosed by a septum.
Their cilia were 4.5-6.0 µm long, however with a diameter of only 150-160 nm. On the exterior of each cilium a tubular system could be detected. Furthermore, the tips of adjacent cilia were interconnected by a kind of fibre bearing a spherical body in its middle. The third type of hair cell (type C), present in the neighbourhood of the second type of hair cell (type B), was chaliceshaped and showed interconnecting fibrils comparable to those found at type B. We believe that these three types of hair cell configurations along with the ganglion cells interconnecting their bases are all components of the gravity organ of the hornet (the Ishay Organ) and together with the cuticular photoreceptors function in the navigation system of the hornet. We further conjecture that the described structures are engulfed by endolymph signals generated by each unit are conducted through neural fibres to the hornet central nervous system.
Keywords : Cilia, hornet, navigation, photoreceptors, SEM/ TEM, noncoating
(Reprinted from: The Journal of Electron Microscopy 48 (1): 63-75 (1999))
Introduction
Social wasps, including hornets (subfamily Vespinae) are insects that build their combs underground, in dim light or complete darkness [1-6]. Comb construction in species of Vespinal is directed towards the gravitational pull of the earth [7,8]. In biological studies it was found that the Oriental hornet is sensitive to a slope of 1.5° and to a delta of radial acceleration as low as 0.1 g [ 9,10]. Suspecting the need of hornets for a gravity sensing apparatus, we have undertaken this study to ascertain the presence and mode of action of such an organ that would enable these insects to sense the direction of the gravitational force during comb building [11]. Indeed we found that on the inner side of the frons plate in social wasps there is a complicated structure composed of static and dynamic nerve fibres, some of which connect between the frons plate and the central nervous system. Encouraged by this finding we conjectured that the interaction between the fibres and the various head structures to which they are attached, is probably part of the proprioceptive system of hornets and is responsible as well for gravity perception. The apparatus involved was accordingly named the "Ishay Organ" and in the same context the presence of cilia on the inner side of the frons plate was also reported [12]. The Oriental hornet Vespa orientalis (Vespinae Hymenoptera) is prevalent in the Middle East, North Africa and West Asia up to India [13]. This hornet's cuticle is predominantly of a brown colour, but two segments of the gaster and several plates on the frons are of a yellow colour due to the presence of symbionts [14]. The frons plate, which bears the gravity organs, is yellow and is built to function as a photovoltaic system [15]. Indeed its upper portion contains parallel transparent plates, similar to these of a solar cell [16]. Additionally there are throughout the cuticle, dispersed several microns apart, distinct pores which are the outlets of peripheral photoreceptors [17]. These peripheral photoreceptors are comprised predominantly of a pore canal and a wide cell underneath the cuticle connected to the nervous system, that are important in hornet orientation and navigation [18]. In the past, several investigators have studied the ciliary structures of different insects mainly by transmission electron microscopy, whereas we have studied hair cells mainly by scanning electron microscopy to obtain a complete 3-dimensional image of the distinct hair cell components involved. Here, we review briefly the related findings:
-The fine structure of the sensory cilium of the auditory receptor of the Australian
cicada (Cyclochila australasiae) has been studied by Young [19]. The
chordotonal sensilla is comprised of one or more bipolar sensory neurons with
the tip of the dendrite enclosed in a specialized structure, the scalopale.
The distal ciliary shaft appears round and bears a ciliary dilatation. A ring
of nine doublet fibrils creates a rod and a tube, the rod bearing a pair of
arms which project towards the neighbouring doublet. The expanded central area
is filled with moderately electrondense amorphous material.
-Heimann [20] found that the first antennae of Conchaecia spinirostris (Ostracoda
Crustacea) display sensory tubes which are arranged either as four tubes with
one seta or as two tubes with three setae. These tubes divide into four regions,
with each tube containing 40-60 ciliated dendrites, some hypodermal and nonneural
cells, and a specialized cuticle. Each dendrite within the tube gives rise to
approximately 25 cilia in a 9 x 2 + 0 pattern. Each cilium splits up into nine
branches which extend to the tip in a partly helical course and form a ring
in the distal part beneath the cuticle. The tubes are covered by a filamentous
surface coat. The ciliary branches probably represent the receptive apparatus,
while the sensory tubes are most likely chemoreceptors.
The coelocapitular sensillum and the antennal hygro and thermoreceptive sensillium of a number of different insects have been extensively studied [21-23]. The cuticular apparatus of the sensillum on the honey bee (Apis mellifera) has a mushroom shaped protrusion devoid of pores and set in a narrow cylindrical pit, positioned in the centre of a shallow depression on the cuticle. Often three or four receptor cells are encountered, three of them bearing unbranched sensory cilia of the 9 x 2 + 0 type and containing densely packed microtubules extending distally into the cuticular apparatus and completely filling its cavity. If there is a fourth receptor it has a thin sensory cilium which terminates beneath the cuticular apparatus, its connecting cilium endowed with armed outer doublets. An outer cavity formed by their enveloping cells is sealed off. The purpose of this scanning electron microscopic investigation was to analyse the 3-dimensional organization of hair cell structures and to provide a detailed classification of the distinct types of hair cells, that form part of the gravity organ of the hornet under investigation.
Materials and methods
Pre-preparation of the hornets.
Hornets, one or two day old, collected from nests in the open field, were
anesthetized with ether and killed thereafter. Subsequently, the heads were
rinsed for 1 min in 0.1 M sodium cacodylate buffer solution and then fixed in
a mixture of 2% GA (glutaraldehyde) and 2% PF (paraformaldehyde) and 0.2% acrolein
in 0.1 M sodiumcacodylate buffer (pH 7.4, 20 °C, for 24 h) [24].
Field emissionscanning electron microscopy.
Specimens were prepared according to the TAO (tannic acid/ arginine/ osmium
tetroxide) noncoating technique, which involved immersion of the samples in
a mixture of arginine-HCl, glycine, sucrose and sodium glutamate [2%, 16 h,
20 °C), rinsing (3x) in distilled water and immersion in a mixture of tannic
acid and guanidineHCI [2 %, 8 h, 20° C], after which the samples were carefully
rinsed (3x) in distilled water. Finally tissues were fixed by immersion in an
OsO 4 solution in distilled water [2%, 8 h, 20 °C], followed by rinsing (3x)
in distilled water, as described previously [25,26]. Dehydration with ethanol
was followed by critical point drying in liquid CO 2 , observations were carried
out with a JEOL FE-SEM, type 6301F, operated at 2-3 kV.
Transmission electron microscopy (TEM).
Small portions of previously observed FE-SEM samples, prepared according
to the GA/ PF/ Acrolein prefixation method and the TAO postfixation procedure,
were carefully orientated, embedded in Epon and ultrathin sectioned. Sections
were poststained with uranyl acetate/ lead citrate and observed in a Philips
TEM, type CM100, operated at 60 kV.
Results
Plate I. An overview of the exterior of the head of the hornet V.
orientalis is shown in Fig. 1; where the left (L) and right (R) compound
eyes as well as the triangular snout bearing the frons plate (fp), coronal suture
(cs) and vertex (vt) are clearly discernible. The ocelli two lateral (a, b)
and one medial (c) are positioned to fit a triangle, with its base orientated
upwards; the two antennae (an) are also observable. A part of the interior of
the head is shown in Fig. 2. Here the crossfractured cuticle (cu) and particularly
the large groups of muscle fibres (mf) are dominant. In the central part of
the figure, groups of ganglion cells (gc) are discernible besides 'brain' tissue
(br), covered on one side by a thin otholitic membrane like layer (om), presumably
a kind of tectorial membrane, damaged at preparation. A large number of nerve
fibres (nf), possibly axons, extend from this area proceeding towards the layer
of hair cells (hc), located on the inside of the cuticle (cu). In a more detailed
image, Fig. 3, closely packed ganglion cells (gc) are found adjacent to groups
of hair cells with cilia. These hair cells represent the type which later is
refered to as hair cell configuration type A, see Figs. 9 and 10.
Nerve fibres (nf) extending from the ganglion cells (gc) penetrate the otholitic
membranelike layer (om), originally covering the hair cells (hc). They interconnect
between the ganglion cells (gc) and the bases of the hair cells as shown at
the left side of figure 3. Another view of the ganglion cells is offered in
Fig. 4. Here, nerve fibres (nf) connect ganglion cells (gc) to the photoreceptors
(pr) inside the cuticle by penetrating through the enveloping otholitic membranelike
layer (om). The small pores (arrows) on the inside of the cuticle (cu) represent
the entrance of the pore canals, each housing a photoreceptor at the endocuticular
region. The axons are directed towards these pores; in the bottom left of the
figure, part of such a photoreceptor (pr) is observable as a spherical body.
A more detailed view of this photoreceptor (pr) is seen in the inset, where
it appears as a woven basket, connected to the top of a kind of lamellar ribbon
(ri). It is obvious that this photoreceptor cell has been damaged, due to forces
exerted at fracturing. The image, nevertheless demonstrates some of the fibrillar
structure of the photoreceptor cell. The substratum (ma) covering the inside
of the cuticle appears as a highly fibrous layer; greater detail of that layer
is shown further on in Fig. 19.
Plate I (for details see text)
Another view of the inside of the head is provided by Fig. 5. On the right of the figure a Plate I (for details see text) lateral (a) and a median (c) ocellus (the second lateral ocellus is obscured by the muscle fibres) can be observed close to the frons plate (fp) and viewed from the rear. The frons plate is covered with compact groups of hair cells, which at this magnification are barely visible; in the bottom left of this figure groups of muscle fibres (mf) can be seen. The somewhat elliptic openings (*) at the bottom left of the figure represent the bases of the two antennae. Both the vertex area adjacent to the bases of the compound eyes (ce) as well as the part close to the three ocelli (a, b, c) are covered with hair cells, as observable in Fig. 6. In more distal parts, the density of hair cells is lower than in the area of the frons plate (fp) and around the compound eyes (ce). The hair cells are arranged in circular groups that are very dense in the area adjacent to the compound eyes but are less dense further up the figure where they aggregate in leafshaped groups, somewhat reminiscent of 'branches'.
Plate II
In the distal area, close to the compound eyes, each more or less circular
group of hair cells (hc) is separated from the adjacent group by a septum (sp),
as shown in Fig. 7. The average diameter of such a group varies between 130
and 300 µm, depending on the location in the head. These groups are similar
to the circular aggregates seen in Fig. 6; however, the density of these groups
within the various areas differs considerably. There are over 20 subunits within
the circular groups, each probably comprising one hair cell unit, with an average
diameter of 30-50 µm. On the right side of the figure remnants of the layer
covering the hair cells can be seen, particularly at the single hair cell to
the right (+). It seems that the covering layer and the septae have a similar
gross morphology. A more detailed image of a few hair cells (hc), here indicated
as hair cell type A, is shown in Fig. 8. This type of hair cell (hc) is surrounded
by a thick and somewhat granular septum (sp) anda large number of stereocilia
to form its characteristic configuration. These stereocilia appear to differ
in length and are grouped together in smaller units, mostly lacking a septum,
as observable in Figs. 9 and 10. More detailed images of the stereocilia occurring
in these units of hair cells are presented in Figs 11, 12 and 13 (FE-SEM) and
in Fig. 14 (TEM image).
A single stereocilium (st) is comprised of a number of tubular structures coated with glycocalyx (*). The average diameter of these tubular structures is appr. 0.5-0.6mm. Figure 13, likewise shows units of (stereo) cilia (st), coated with glycocalyx (*). Many of them (~ 15-20) are arranged in a bundle or unit; a nerve fibre (nf) is seen alongside the cilia (st). This type A hair cell is similar to the one shown in Fig. 3 adjacent to the ganglion cells. Remnants of glycocalyx or haemolymph obscure the view, yet the entire aggregate is clearly a large cluster of stereocilia whose diameter ranges between 7 and 8 µm. Figure 14 represents a TEM image of an ultrathin section of a few stereocilia (st) with an average diameter of 500-600 nm, comparable to what has been shown in Fig. 12. Stereocilium type A is composed of nine tubular structures, separated from one another by a rather thin layer of electron dense material, probably glycocalyx (*), heavily stained as result of the tannic acid/ arginin/ osmium tetroxide (TAO) noncoating method. At the bottom of the picture a sidelink (sl), bridging between two adjacent stereocilia, is observable. In a few other places sidelinks or places where they were present before the sectioning are observable, although less clearly.
Plate III
Particularly in the vicinity of a peripheral photoreceptor pore canal (po)
outlet, cilia can be seen which are arranged in groups forming a type of hair
cell configuration, further on indicated as hair cell type B. However, these
hair cells are not or are only partially surrounded by a septum (sp), as can
be observed in Fig. 15. The cilia seem to arise directly from a highly fibrillar
matrix (ma) covering the multilayered cuticle (compare with Fig. 4). The average
diameter of hair cell type B is appr. 13 µm, which is thus larger than the diameter
of the ones seen in Fig. 12, although smaller than the diameter of those shown
in Figs 8 and 9. Hair cell type B is shown in more detail in Fig. 16, which
also shows the presence of a septum (sp) at one side and the occurrence of (stereo)
cilia (st), emerging from the cuticular substratum (ma). The latter cilia are
clearly longer and thinner (average diameter appr. 150 nm) than those shown
previously.
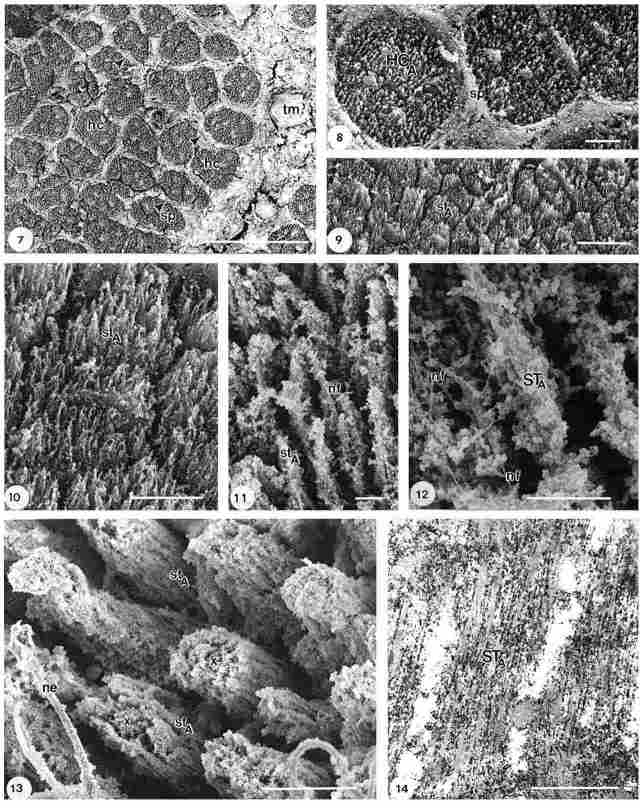
Legends of Plate II. (Fig. 7) Larger unit with circular groups of hair
cells (hc) type A with stereocilia, separated from one another by septae (sp).
At the right, one hair cell (+) still is covered with an otholic membranelike
layer. Bar = 100 µm. (Fig. 8). A few circularly shaped units of hair cells (he),
separated by a septum (sp), with clusters of stereocilia. Bar = 10 µm. (Figs.
9, 10) Subdivision of part of one hair cell of type A with stereocilia (st)
of different length, no septum present. Bar = 10 µm. (Figs. 11, 12) Detailed
image of a stereocilium (st) of type A with neurofibrils (nf) inside, note glycocalyx
(*) at in and outside stereocilia. Bar = 1 µm. (Fig. 13) Subdivision of a hair
cell (hc) type A, consisting of a group of appr. 20 stereocilia (st) around
a central area. Bar = 10 µm. (Fig. 14) TEM longitudinal section of a few stereocilia
of type A hair cell, the middle one with a width of appr. 500 nm, consisting
of about 9 closely packed tubular structures, interspaced by heavily stained
glycocalyx (*). Note sidelink (sl) between two adjacent stereocilia. Bar = 500
nm.
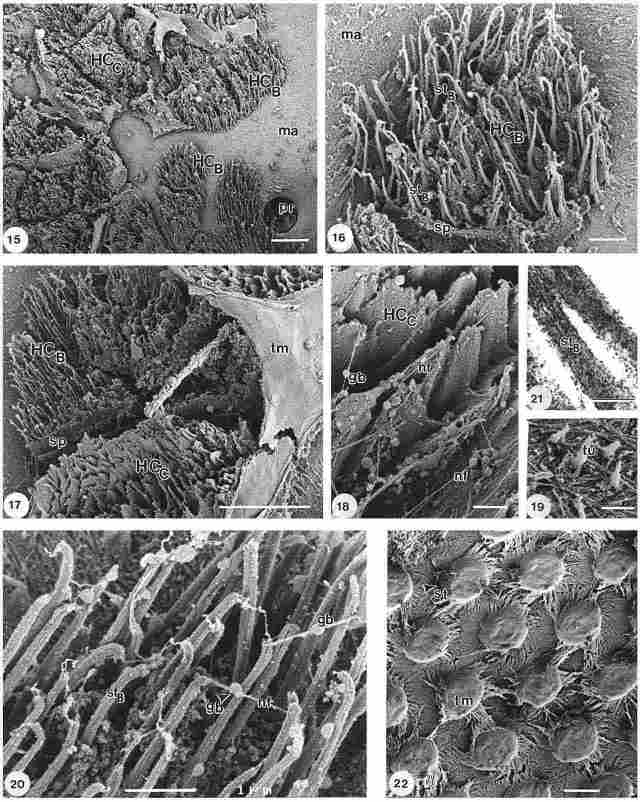
Legends of Plate III. (Fig. 15) Type B hair cells (hc) with cilia adjacent to
pore-canal entrance (po) of photoreceptor cell, partly enclosed by a septum
(sp); note matrix material (ma) covering the cuticle inside wall. Bar = 10 µm.
(Fig. 16) Detail of type B hair cell (hc), with cilia (st) emerging from fibrillar
matrix (ma), part of the structure is bordered by a septum (sp). Bar = 2mm.
(Fig. 17) Type B hair cell (hc) with cilia adjacent to chalice structure (ch)
and otholithic membrane (om), part of type B and typeC hair cells are bordered
by a septum (sp). Bar = 10mm. (Fig. 18) Detail of chalice type C hair cell (hc)
with fibrillike connections (fi) with a globular body (gb) in the middle. Bar
= 1 µm. (Fig. 19) Detail of fibrillar covering of inside of cuticle with tufts
(tu). Bar = 1 µm. (Fig. 20) Detail of stereocilia (st) of type B hair cell with
fibrillike connections (fi) bearing a globular body (gb) at the middle; note
longitudinal lines at cilia and the presence of glycocalyx material (*). Bar
= 1 µm. (Fig. 21) TEM section of stereocilia (st) of type B hair cell, note
rather compact structure. Bar = 200 nm. (Fig. 22) Hair cells at distal part
of vertex, covered with a slightly inferiorly turned lamellar layer (la), at
some places showing part of the hair cell (hc) underneath. Bar = 10µm.
Figure 17 represents a higher magnification of an area similar to that shown in Fig. 15. Hair cell type B seen here is adjacent to a structure resembling a chalice (ch), this is the third type of hair cell configuration, further on indicated with hair cell type C. Remnants of an otholitic membranelike covering layer (om) are seen at the right part of the figure. It could have a similar function as the tectorial membrane in mammals. A rather granular septum (sp), abutting on one side of hair cell type B, is also observable. Frequently hair cell type B and C are encountered in the very same unit as witnessed at the top left of this figure. A detailed view of hair cell type C is provided in Figure18. Here the granularity of the structure and the presence of very thin fibrils (fi), within the middle a globular body (gb), connecting the chalice (ch), are clearly seen. A detailed image of the fibrillar substrate lining the inside of the cuticle, is provided in Figure 19, showing tufts (tu), which have a larger diameter at their base ( ~ 0.25mm) , at the top (~ 0. 15mm). A higher magnification of the (stereocilia of the of hair cell type B is shown in Figure 20. Here, individual cilia (st) display a certain pattern of lines along their length. The cilia are interconnected at their top by a thin fibril, protruding from the centre of the stereocilium at the top, bearing a globular extension (gb) in the middle. In between the individual cilia a granular substance is found, which could represent glycocalyx or endolymph material. The individual stereocilium base has an average diameter of about 140-150 nm, while its length varies between 4.6 and 6.0 µm. Figure 21 shows a TEM detail of a stereocilium (st) of hair cell type B. Comparing the stereocilia of a type B hair cell (fig. 21) with those of type A hair cell (Fig. 14), it is obvious that the diameter of type A is ~ 500-600 nm, while the diameter of type B is only ~ 120-130 nm, as calculated from TEM images. This is in agreement with the SEM results, as based on the visualization of stereocilia after tannic acid/ arginin/ osmium tetroxide treatment, which often produces larger complexes with osmium. One stereocilium of type A hair cell consists of about 9 subunits held together by glycococalyx-like material, heavily stained by the TAO noncoating method. Each subunit of it is ~ 35-40 nm in width. A cilium of hair cell type B does not show much of a subdivision, it is heavely stained as result of the TAO treatment, giving it a somewhat coarse appearance. In the longitudinal section dark patches are seen; sometimes a thin enveloping heavely stained layer is seen, giving the impression of a rather compact unit embedded in glycocalyxlike material. The thin lines observable at the surface of the stereocilia type B (Fig. 20) suggest, together with Fig. 21, that the lining appears at the outside surface of the stereocilium.
Figure 22 presents hair cell units (hc) at the distal part of the vertex, each enclosed like a box by a thin striated lamellar layer (la). The layer has been somewhat damaged by the drying procedure used, causing shrinkage and resulting in deformation of the layer and of the caplike structure at the tip of each hair cell. At one point the presence of stereocilia underneath this layer is also discernible. It is possible that this layer represents the tectorial membrane-like structure seen before.
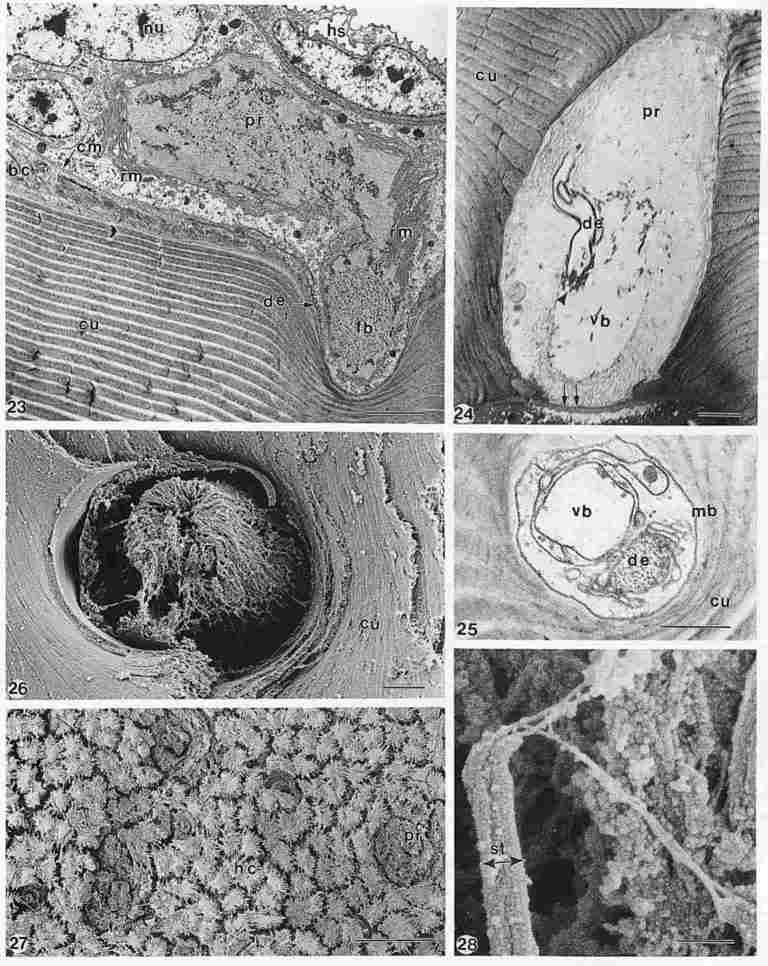
Legends of Plate IV. (Fig. 23) Longitudinal TEM section of photoreceptor cell
(pr) with nucleus (nu) inside the multilayered cuticle (cu). The two compartments
of the cell are partly filled with fibrillar material (fb) and enclosed by a
folded outer rhabdomer membrane (rm) containing retinoides. Basal cuboidal cells
(bc) extend along the inside of the cuticle, covered by the cell membrane (cm)
of the photoreceptor cell (pr). A dendrite structure (de) entering the photoreceptor
cell and typical haemolymph spaces (hs) can also be observed. Bar = 2 µm. (Fig.
24) Other TEM section of photoreceptor cell (pr), almost perpendicular to that
of Fig. 23, is seen inside the cuticle (cu). A dendrite-like structure (de)
is entering the cell close to the vitreous body (vb). A membrane(íí),
closing off the pore canal from the photoreceptor area, serves as an aperture
to light from the outside. Bar = 2mm. (Fig. 25) Other TEM ultrathin section
of a photoreceptor cell inside cuticle (cu), showing the vitreous body (vb),
a dendrite (de) enclosed by various membranes (mb) and the spot where synaptic
processes take place (Y). Bar = 1 µm. (Fig. 26) FE-SEM image of a photoreceptor
cell, cytosk-eleton consist of a fine network of fibres with a spool-like appearance
extending above cuticle (cu); image compa-rable with Fig. 4 (inset). Bar = 2
µm. (Fig. 27) FE-SEM image of inside area of the head covered by hair cells
(hc) with cilia surrounding photoreceptor cells (pr) at pore canal openings.
Bar = 50 µm. (Fig. 28) Detailed FE-SEM image of apical side of stereocilia of
type B hair cell, note several kinocilia-like extensions at the top of cilium
connected to other stereocilia. Bar = 200 nm.
Plate IV
Figure 23 shows an almost longitudinal section of a photoreceptor cell,
located inside the cuticle (cu) extending to the left and the right. The cell
nucleus (nu) and the basal membrane (cm) covering the inside of the cuticle
with the basal cells (bc) are well observable, so are the spaces (hs) with endolymph
at the top of the image. The photoreceptor cell seems to consist of two compartments
containing fibrillar material (fb), separated by a membrane. The photoreceptor
cell is enclosed by a rhabdomerer membrane (rm) appearing as a folded membrane
containing retinoides. A dendritic fibre (de) enters the photoreceptor cells
at the left side of the cell close to the cuticle (cu). The thickness of the
cuticular layers is clearly increasing towards the outside cuticle. Figure 24
presents another view onto the photoreceptor cell (pr) enclosed by the cuticular
layers (cu). It consist of different compartments, some filled with fibrillar
material some are almost empty forming the vitreous body (vb) of the cell. A
dendrite (de) leaving the photoreceptor cell and the location of the sinaps
processes (m) can be observed. The image has been orientated in the same way
as Fig. 23, at the bottom of the picture a membrane is found, closing off the
photoreceptor area from the pore canal, serving as a kind of aperture for light
coming in. The space below the membrane, the pore canal, is filled by resin
as a result of the embedding procedure; the white spots are electron beam irradiation
artefacts. Figure 25 represents another section of the photoreceptor cell orientated
almost perpendicular to that of Fig. 23, the presence of a vitreous body compartment
(vb) and a dendrite (de) is well observable, obviously they are surrounded by
membranous structures (mb). Figure 26 represents a FE-SEM image of a photoreceptor
cell( pr), a rather fibrillar structure assuming a sort of spool shape forming
the cytoskeleton of the cell. The photoreceptor cell is closely surrounded by
the cuticular layers (cu), it appears that the cell is surrounded by a membrane
(R) as observed in the TEM images before. The image shows some similarity with
the inset of Fig. 4 and on the other hand with the fibrillar material found
in the compartments observed in the TEM images of Figs. 23, 24 and 25. As shown
previously (Fig. 15), pore canal outlets with photoreceptor cells inside are
often surrounded by stereocilia of type B hair cells. In Figure 27, that part
of the photoreceptor cells (pr) extending outside the cuticle, similar in Fig.
23 is visible, surrounded by hair cells (hc) with cilia. Obviously some drying
artefacts have occurred, both at the photoreceptor cells and the hair cells,
causing shrinkage and aggregation of cilia. Due to the location of the photoreceptor
cell inside the cuticle, optimal preservation of the delicate structures is
rather difficult, which accounts for the various appearances of the cell particularly
in FE-SEM and to some extent also in TEM. The variance in appearances in TEM
though is mainly the result of different cutting faces obtained. Finally, Figure
28 represents a higher magnification of the apical side of a stereocilium of
type B hair cell, at least three kinociliar extensions are found, which connect
with other stereocilia; due to fixation and drying forces exerted, their shape
might be altered slightly.
Discussion
The hair cell is a receptor cell in a sensory system. It performs an essential
function, namely, of transducing a mechanical sensory input into electrical
signals [27]. It is thus a mechanoelectric transducer element which is vital
to all functional properties of vestibular organs [26]. The name hair cell for
the structures found, is chosen based on their resemblance to hair cell found
in the organ of Corti as part of the cochlea of mammals, though, the origin
of the cilia of the hair cells in the hornet are different from those in the
organ of Corti. Those of the hornet seems to have their origin at the highly
fibrous substrate covering the inside layer of the cuticle, nevertheless, the
arrangement of various cilia form specific structures. We conjecture that hair
cells act as mechano receptors to gravity dependent accelerations. The hair
cells are encountered in the head of the hornet covering almost all the area
of the frons plate and vertex, i. e. the areas not occupied by muscle insertions,
peripheral photoreceptor pores and the three ocelli. The overall covering with
hair cells with a large number of cilia is enhancing the sensitivity to detect
the smallest directional changes, in particular in the dark. The hair cells
are connected to ganglion cells and to the nerve fibres in the Ishay Organ which
ultimately enter the brain. Why have the described hair cells remained uninvestigated
until now? To our knowledge the ultrastructure of these ciliabearing cells located
in the interior of the head cuticle close to the frons and the ocelli has never
been studied extensively by SEM or TEM. There are a number of reasons for that.
To begin with, standard GA/ PF prefixation and 0sO 4 postfixation do not preserve
these structures adequately, moreover, there is leaking out of certain components,
the production of artefacts due to irradiation damage and inevitable shrinkage
as a result of the dehydration/ drying process. Secondly, the need for relatively
high acceleration voltages (15-25 kV) used in the past, required the use of
relatively thick (8-10 nm) conductive layers to avoid charging. These layers
are often too thick and granular, obscuring surface details [29]. Furthermore,
structures may be obscured or masked by the high ratio of secondary electrons
generated within the investigated structures as compared to the secondary electrons
generated at the very surface of the structures. Thirdly, a combination of SEM
and TEM procedures on the same specimen, is virtually possible only on optimally
preserved structures processed initially for SEM and subsequently embedded for
TEM. [30- 32]. The use of a GA/ PF/ acrolein prefixation followed by tannic
acid/ arginin/ osmium tetroxide noncoating postfixation provides optimal preservation
of structures containing glycoproteins and mucopolysaccharides [33- 35]. The
use of a low accelerating voltage (23 kV) in FE-SEM offers high resolution and
better surface topographic imaging, thus contributing to an improved image of
ciliary structures. The low brightness of standard W or LaB6 electron sources
is unsufficient to allow low kV SEM operation at an acceptable resolution. Descriptive
review of the encountered hair cells and photoreceptors. Three different types
(A, B, C) of hair cells or hair celllike structures could be distinguished within
the head of V. orientalis. Hair cell type A is found in the frons and
vertex close to the ocelli and are arranged in more or less round clusters having
a diameter of 180-300 µm, the clusters separated from one another by a septum
(Fig. 6). These roughly round clusters were in turn organized in larger aggregates
which assumed either an angular shape and high density when located closed to
a compound eye or a leaflike shape and a lower density when located in the periphery.
Within each such aggregate (Fig. 7) there were between 20 and 25 clusters of
hair cells with cilia, each cluster averaging 30-50 µm in diameter and separated
from one another by septae. In turn, such hair cell clusters subdivided into
a large number of subclusters having a diameter of ~7-8 µm. The cilia in such
a subunit vary in length as evident from the light charging of the tips of the
individual cilia. These subclusters are discrete within the hair cell cluster.
At higher magnification such a subcluster reveals a central area of app. 3-5
µm in diameter, surrounded by a ring of single stereocilia. Each stereocilium
has an average width of 130-140 nm and seems slightly thicker at the base than
at the top. The length of the single stereocilium in a given subcluster is app.
12.5 -14 µm.
In mammals, the length of the stereocilia in the outermost row of hair cells in the organ of Corti is ~ 2.5-3.8 µm, so they are considerably shorter than these of the hornet [37]. At this type of cilia, sidelinks have been observed, probably involved in the process of cilia activation and inhibition. Most of the exterior of the cilia is covered with a somewhat granular material deriving either from the glycocalyx or the endolymph, probably consisting of glycolipids, glycoproteins and other materials. The anionic sites of these compounds react with cations such as tannic acid and osmium. In SEM imaging subunits of a single cilium can be discerned, namely, microtubules whose diameter is ~ 30-40 nm. This is in agreement with the data obtained from inspection of a section of a single cilium in TEM where the diameter was ~ 40 nm. Hair cell type B possesses stereocilia which are close to the outlet of the photoreceptor at the cuticular pore canal and are arranged in groups, but lack a septum or have only a partial septum. The number of stereocilia within a type B hair cell is smaller than in type A and moreover, the diameter of the type B cell is ~ 4.6-6.5 µm as compared to a diameter of 20- 50 µm for type A. The stereocilia of hair cell type B are not only different in length from type A, but are also much smaller in diameter (150-160 nm) as compared to 500-600 nm. In some places, the stereocilia just seem to be an extension of the fibrous matrix coating the inside of the cuticle. Such 'single' cilia are often connected at their very top to adjacent cilia by means of a thin fibril bearing a globular swelling in its middle. Some of the stereocilia have more than one kinociliar extension The length of these interconnecting fibrils is ~ 1 µm. It is possible that the globular swelling is connected to the base of the tectorial membrane covering the cilia, in which case it could be similar to the bulbs found on nonmotile kinocilia that are connected to the otolithic membrane in mammals [36]. They certainly differ from the tip and side links encountered on stereocilia of the hair cells of the outer rows in the organ of Corti in mammals, which contribute to mechanosensitivity of the hair cells [37-41]. In the organ of Corti of mammals [36-41] in the W-shaped stereocilia cross connections occur either between the stereocilia in the same row or between short and medium length stereocilia. Their function is to mitigate the effect of excessively strong vibrations on single stereocilia. Possibly the septae between groups of cilia in V. orientalis have a comparable function. The variation in both the density and the shape of the cilia may perhaps reflect on their particular physiological function in the gravitic system of the hornet. Hair cell type C is a structure which until now has not yet been explained; they are the chaliceshaped units located close to the groups of cilia surrounded by a septum (conceivably representing the bases of muscles). These structures seem to be virtually empty shells, although transsected by very fine fibrils and bearing very small globular encrustations at their bottom. The material from which these structures are composed seems to be similar to that of the septae. The fact that these structures are often found devoid of content could be an artefact, that is, they may have originally been filled with endolymph which was leached out during the rinsing step in the preservation procedure. The original contents, if any, could perhaps play a role in the gravity system, conceivably slowing down excessively rapid movements. These peculiar structures are mostly located close to the type B hair cells, often separated from them by a septum. Hair cells of type B and the type C structure are frequently covered by the so called tectorial membrane as is well demonstrated in Fig. 17. Photoreceptor cells are located in the multilayered cuticle, very much enclosed in a pore canal formed by the cuticular layers, and are found in the head and in the abdomen at the yellow strip. The ones presented here are found in the head and are therefore rather difficult to reach for fixation mixtures, resulting in an uneven preservation of the various structures making up the photoreceptor cell. The comparison and interpretation of FE-SEM and TEM photoreceptor images for the same reason is rather difficult. The presence of fibrillar structures arranged in a kind of spoolshape seems obvious from both FE-SEM and TEM images. The FE-SEM specimens of photoreceptors could be slightly damaged, because they are obtained after fracturing the cuticle which often leads to the excertion of pulling forces. The light enters the pore canal at the epicuticle side at a certain angle, due to the shape of the eaves at the outside of the cuticle, and is transported to the photoreceptor cell. A membranous structure acts as a kind of aperture to restrict the amount of light entering the photoreceptor cell. The light is transformed into a neural impulse which is transferred to the ganglion cells, the presence of a kind of nerve connection is acknowledged both in FE-SEM and TEM. Due to the special light entrance conditions, small directional changes of the body can be detected by the photoreceptor cell found in the cuticular layers of the head and the abdominal area at the yellow stripe. Clearly this is only possible in daylight; navigation in the dim is carried out most probably by the three ocelli. A thin otholitic membrane-like sheet covers the cells and suggests a certain similarity with the tectorial membrane, covering the longest stereocilia of the outer hair cells of mammals. The membrane-like sheet comes in contact with the tips of the cilia, on this membrane some small otholites can be found, Thet can be compared with maculae of the vertebrate vestibular system. This analogy suggests that the role of the structure is in the sensation of linear accelerations. Over the tectorial like membrane at the inner side, a large number of ganglion cells can be seen. Some dendrites connect these cells to the photoreceptors, others connect them to different types of ciliary cells interconnecting between ganglion cells. This structure suggests that the role of these ganglion cells is to integrate information coming from the ciliary cells of the total area of the head covered by these cells. This information is further integrated with information coming from the photoreceptors of the same area. Gravity direction and light are the cues for hornets navigation and so this is probably the ganglion of navigation.
Conclusions
We have encountered three types of structures in the head cuticle of the
Oriental hornet, all of them naturally covered by an otholitic membrane-like
sheet with small crystallike structures, possibly otoliths. Not in all cases
this sheet is well observable in the images shown, because some kind of disruption
takes place as a result of drying forces. A similar process is seen at the tectorial
membrane covering the rows of hair cells in the organ of Corti in mammals, where
the tops of the longest stereocilia of the outer rows of hair cells separate
from the tectorial membrane undersurface at appropriate fixation of the organ
of Corti, if not the stereocilia can fracture at their base [31]. The mutual
proximity of these three structures and of the common ganglion cells, suggest
that they are closely related. The cilia cells of the frons probably are gravireceptors.
The type A hair cells far outnumber the type B cells. The latter are mostly
found close to the pore canal outlets of the peripheral photoreceptor. All these
structures are located on plates in the anterior upper portion of the head,
exterior to the nerve fibres of the Ishay Organ, but in close contact with them.
Type A stereocilia are longer and have a larger diameter than type B. In crosssection
type A stereocilia display tubular structures, particularly in TEM images, and
also thin fibrils. Type B stereocilia do not show fibrils, although they have
a tubular substructure visible from the outside. Long extensions acts as a kind
of kinocilium which interconnects adjacent stereocilia. The small spherical
structure in the middle of such a kinocilium touches the covering otholitic
membrane and is similar to the vestibular kinocilia in mammals. From the scanty
endolymph still retained between the stereocilia, particularly of type B, it
is reasonable to assume that the rinsing steps probably removed a great deal
of the original amount. This endolymph or hemolymph probably acts in the same
way as it does in mammals in the hair cells of the vestibular area. Both type
A and type B cilia seem to originate fro the highly fibrous substrate covering
the cuticle on the inside. Indeed small tufts are seen to emerge from the fibrillar
coat, probably indicating the incipience of stereo or kinocilia. Whether the
chalice structure (type C), which sometimes is surrounded by cilia of type B
hair cells, has a special function when filled with endolymph remains to be
ascertained. The presence of all kinds of rather small spherical units and axonfibre
connections in these shelllike structures suggests for them a role in hornet
navigation. If this be true, then the slightest movement of endolymph within
this structure could possibly generate a signal detectable by the fibrils connecting
the various units within it.
An important question that remains to be answered is whether the cilia are stereo-or kinocilia. The cilia which are part of the hair cells in the organ of Corti in mammals are stereocilia, apart from their role as sensory cilia. Although the cilia under investigation bear a certain resemblance to the microvilli of the intestinal epithelium and to cilia in lung epithelium tissue, they have a central core which is ~ 3.5 µm in diameter and surrounded by 7-8 tubular structures. According to Ham and Cormack in their texbook on Histology [42], cilia are hairlike processes extending from free surfaces of unicellular organisms and cells in the body which are about 5-15 µm long and ~ 200 nm in diameter. They are developed from centrioles, whose wall is composed of nine longitudinally disposed and parallel bundles of microtubules, each bundle containing three microtubules (triplets). The triplets of a centriole are held in position by fibrillar material together forming the wall of a cylinder. Cilia can undergo extensive modification, as occurs in the receptor cells of organs of special sense. These modified cilia become very important parts of the arrangement by which nerve impulses are initiated as result of exposure to certain forms of energy, such as the rods and cones of the eye, representing modified cilia which are receptive to light energy. Stereocilia are tufts of extremely long microvilli projecting towards the lumen from the free surface of the (tall) cell. Mostly no characteristic pinocytotic vacuoles are shown in the apical cytoplasm of those cells. The free surface of a hair cell has hairlike processes extending from it, mostly called stereocilia. Stereocilia are narrow towards the origin and widen towards their tip. Vibrations predominate in a given region of the organ of Corti vibrating at this particular frequency and is sensed by its hair cells as a result of their microvilli. They get displaced with respect to the tectorial membrane in which their tips are embedded. This causes the hair cells to alter the pattern of impulse activity in the afferent branch of the acoustic nerve they contact. Probably some large efferent nerve endings that contain synaptic vesicles are believed to pass impulses to the hair cells. From the above presented data it seems likely that the cilia under investigation are stereocilia.
References
1. Ishay J S, BytinskiSalz H, and Shulov A (1967). Contributions to
the bionomics of the Oriental hornet vespa orientalis. J. Entontol. 2: 45-106.
2. Wilson E 0 (1971). The Insect Society, (The Belknap Press of Harvard University
Press, Cambridge, USA).
3. G–epes G (1972). Les Guikia Sociales, (Hymenoptera vespidae), (Masson et
Cie, Paris).
4. Spradbery J P (1973) Wasps. In: An Account of the Biology and Natural History
of Solitary and Social Wasps, ed. Spradbery J P, p. 480, (Sidgwick and Jackson,
London).
5. Edwards R (1980). Social Wasps. Their Biology and Control, (Rentokil Lid,
East Grinstead, UK).
6. Matsuura M and Yamana S (1990). In: Biology of Vespine Wasps, eds. Matsurna
M and Vamane S, p. 323, (Springer Verlag, Berlin).
7. Ishay J S and Sadeh D (1975). Direction finding of hornets under gravitational
and centrifugal forces. Science 190: 802-804.
8. Ishay J S and Sadeh D (1977). Geotropism of hornet comb construction under
persistent acceleration. Beh. Ecol. Sociobiol. 2: 119-129.
9. Ishay J S, Rosenzweig E, and Abir I (1986). Geotropic sensitivity exhibited
by single hornets: the influence of cast, age, light and temperature. Adv. Space
Res. 6: 37-40.
10. Ishay J S, Rosenzweig E, Rosenzweig O, and Berke S (1989). Gestropic sensitivity
of hornets. Adv. Space Res. 9: 147-155.
11. Arcan L and Ishay J S (1993). Internal micromorphology of the frons plate
in females in the Oriental hornet. J. Morphol. 217: 147-159.
12. Ishay J S, Landsberg A and Pelah S (1996). Micromophology of the fibers
behind the frons plate and its adjacent regions in the Oriental hornet (Hymenoptera,
Vespinaes. Scann. Microsc. 10: 187-206.
13. Bodenheimer K (1997). Uber die Aktivitat von Vespa orientalis F. im Jahresverlauf
im Palastina. Zool. Anz. Leipzig C11: 135-140.
14. Ishay J S and Shmuelson M (1994) Symbiosis with a fungus produces the colored
stripes in social wasps. Physiol. Chem. Phys. & Med. NMR 26: 245-260.
15. Ishay J S, Goldstein 0, Rosenzweig E, Kalicharan D and Jongebloed W L (1997).
Hornet yellow cuticle microstructure: A photovoltaic system. Physiol. Chem.
Phys. & Med. NMR 29: 71-93.
16. Maycock P D and Stirewall E N (1981). Photovoltaics, Sunlight to Electricity
in One step, (Birch Horse Co., Andover, MA).
17. Goldstein O and Ishay J S (1996). Morphology of a putative new peripheral
photoreceptor in social wasps. Physiol. Chem. Phys. & Med. NMR 28: 129-136.
18. Jongebloed W L, Kalicharan D, Goldstein 0, Rozenzweig E, Want JJ L van der,
and Ishay J S (1997) The microstructure of the photoreceptor and yellow strip
of the hornet Vespa orientalis I: FE--SEM and TEMobservations. In: Jaarboek
NVVM 1997, eds. Koerten HK, Drost G and Berendsen W, pp 91-94, (Press Point,
Rijnsburg, The Netherlands).
19. Young D (1973) The fine structure of the sensory cilium of an insect auditory
receptor. J. Neurocytol. 2: 47-58.
20. Heimann P (1979) Finestructure of sensory tubes on the antennule of Chonchoecia
spinirostris (Ostracoda, Crustacea). Cell Tiss. Res. 202: 461-477.
21. Lacher V( 1964). Elektrophysiologische Untersuchungen an einzelnen Rezeptoren
fur Geruch, Kohlendioxide, Luftfeuchtigkeit und Temperatur auf den Antennen
der
22. Altner H and Prillinger L (1980) Ultrastructure of invertebrate chemo--,
thermo, and hygro-receptors and its functional significance. Int. Rev. Cytol.
67: 69-139.
23. Yokohari E (1983) The coelocapitular sensillum. An antennal hygro--and thermoreceptive
sensillum of the honey bee, Apis mellifera L. Cell Tiss. Res. 233: 355-365.
24. Jongebloed WL, Kalicharan D, Zhang T and Rawson DM (1997). The zebrafish
(Brachydanio rerio) embryo egg inner and outer membrane, a FESEM (I) and TEM
(II) investigation. In: Jaarboek NVvM 1997, eds Koerten HK, Drost G, Berendsen
W, pp. 94-97, (Press Point, Rijnsburg, The Netherlands).
25. Jongebloed W L and Kalicharan D (1994) Tannic acid/ arginine/ osmium tetroxide
fixation of rat tissue by the microwave procedure. Beitr. Elektronenmikroskop
Direktabb Oberfl. 27: 243-252.
26. Jongebloed W L, Dunnebier E A, Kalicharan D, and Albers F W J (1996). Demonstration
of the fine structure of stereocilia in the organ of Corti of the guinea pig
by field emission scanning electron microscopy. Scann. Microsc. 10: 147-164.
27. Hudspeth A J (1985) The cellular basis of hearing: The biophysics of hair
cells. Science 230: 747-752. 28. Anniko T (1984) Cytodifferentiation of cochlear
hair cells. Am. J. Otolaryng. 4: 375-388.
29. Stokroos I, Kalicharan D, and Jongebloed W L (1995). A comparative study
of ultrathin coatings for high resolution FEGSEM. Beitr. Elektronenmikroskop.
Direktabb. Oberfl. 28: 165-172.
30. Jongebloed W L, Kalicharan D, Vissink A, and Konings A T W (1992). Application
of the OTOTO noncoating technique; A comparison of LM, TEM, and SEM. Microsc.
& Anal. 28: 3133.
31. Jongebloed W L, Kalicharan D, and Worst J G F (1994). High resolution FEGSEM
im ages of biological tissue. Proc. 13th Int. Conf. on EM (ICEM), Paris. Vol
3a: 777 778.
32. Jongebloed W L and Kalicharan D (1996). Low voltage visualization of glycocalyx
(like) structures in various biological tissues. Beitr. Elektronenmikroskop.
Direktabb. Oberfl. 29: 201-212.
33. Murakami T (1974). A revised method for uncoated SEM specimens. Arch. Hist.
Jap. 36: 189-193. 34. Murphy J A (1978). Noncoating techniques to render biological
specimens conducive. Scann. Microsc. (ITTRI), Chicago: 175-195.
35. Chaplin A J (1985). Tannic acid in histology, a historical perspective.
Stain Technol. 60: 219-231.
36. Kessel RG and Kardon R H (1979). Tissues and Organs, A Text Atlas of Scanning
Elec tron Microscopy . (WH Freeman and Co, San Francisco).
37. Pickles JO, Comis SN, and Osborne M P (1984). Cross links between stereocilia
in the guinea pig organ of Corti and their possible relation to sensory transduction.
Hear. Res. 15: 103-112.
38. Furness D N and Hackney C M (1985). Crosslinks between stereocilia in the
guinea pig cochlea. Hear. Res. 18: 177-188.
39. Hackney C M and Furness D N (1986). Intercellular crosslinkages between
the stereociliary bundles of adjacent hair cells in the guinea pig cochlea.
Cell Tiss. Res. 245: 685-688.
40. Osborne M P Comis SD and Pickles JO (1988). Further observations on the
fine structure of tip links between stereocilia of the guinea pig cochlea. Hear
Res 35: 99-108.
41. Van Benthem P P G, De Groot J C M J, Albers F W J Veldman J E, and Huizing
E H (1993). Structure and composition of stereocilia crosslinks in normal and
hydrophic cochleas of the guinea pig. Eur Arch. Otorhinolaryngol. 250: 73-77.
42. Ham A W and Cormack D H (1974). Textbook of Histology 8th ed. , Ch 28: The
eye and the ear, pp. 903-940, JB Lippincon Co, Philadelphia).