Altered behaviour in hamsters conceived and born in hypergravity H.N.P.M. Sondag, H.A.A. de Jong, W.J. Oosterveld
Abstract
We studied vestibular function in 36 adult
hamsters (1 month old) conceived and born in either hypergravity (n=21) or normal
gravity (n=16). Four groups were made: a) HL group (n=13): 20 weeks in 2.5 G
and 14 weeks in 1 G, b) HS group (n=8): 4 weeks in 2.5 G and 30 weeks in 1 G,
c) CON group (n=9): 34 weeks in 1 G and d) ROT group (n=7): 20 weeks in rotation
in 1 G, located at the center of the centrifuge and 14 weeks 1 G. We investigated
locomotion, swimming, air-righting and the susceptibility to rotations during
this period. We found that HL and HS hamsters had no disturbances in gait and
locomotion but that their swimming ability (drowning, circling and speed of
swimming) was disturbed. Furthermore, the HL hamsters showed less treadmill
activity during 2.5 G, had balance disturbances, showed less rearing activity
in the open field, were less susceptible to rotatory accelerations and showed
fewer correct air-rightings than the other groups. After 3 months of normal
gravity, these differences between groups disappeared except for the disturbed
swimming ability and the number of correct air-righting responses. The hamsters
subjected to rotation without the increased G-load (ROT group) had balance difficulties
which can partly be attributed to their higher bodyweight.
We conclude that the structural alteration to hypergravity in the otoconial
layers of the vestibular end organs that we described in another experiment
is one of the causes for the behavioural disturbances found in hamsters prenatally
developed in hypergravity. Furthermore, hypergravity and rotation have a different
effect on vestibular evoked behaviour.
Keywords: centrifugation, locomotion, balancing, air-righting, swimming, otoconia.
Introduction
During space shuttle missions more than
60% of the astronauts complained about symptoms of motion sickness (Space Adaptation
Syndrome) which decreases their performance during flight. It is suggested that
conflicting sensory input from vestibular and visual perception system causes
these symptoms (Homick, 1977; Reschke, 1986; Paloski, 1993). Because it is difficult
to conceive animals in space and test their perceptive-motor skills for a long
period in different gravity levels, we breed animals in a hypergravity environment.
In a centrifuge it is possible to subject animals to hypergravity for long periods
of time without severe effects on their anatomy and their well-being (Oyama
and Zeitman, 1967; Economos et al. 1982). Furthermore, these experiments give
us the opportunity to study the behaviour of animals during hypergravity as
well as during normal gravity but also to assess possible structural alterations
in physiological systems due to the hypergravity environment.
In the experiment we present here, hamsters were conceived and born in a centrifuge
under hypergravity conditions (2.5 times normal gravity, 2.5 G). In a previous
article we reported a structural alteration in the otolith organs of hamsters
developed in hypergravity conditions, i.e. changes in the relative size of areas
with large, medium-sized or small otoconia and the development of these areas
on the utricular patch (Sondag et al. 1996b). Here, we report the effects of
hypergravity on the behaviour of these hamsters. During hypergravity, their
locomotor activity will be assessed by means of an activity wheel. Locomotion
of these hypergravity developed hamsters might be altered when subjected to
normal gravity because of changes in neuromuscular functions. Also tests which
assessed the vestibular function, swimming and balancing were included in our
experiment. was investigated with tests
Material and methods
The study was done in a group of 36 subadult golden hamsters (Mesocricetus auratus). The hamsters were conceived born at our laboratory (parent generation came from Harlan, Zeist, The Netherlands). Of this group, 21 hamsters were conceived and born under conditions of 2.5 G (HG hamsters) in the centrifuge, while the other 16 were conceived and born in normal gravity (CON hamsters). A number of animals of the HG group were placed in normal gravity after weaning (HS for Short Hypergravity period) to investigate whether age is an important factor concerning adaptation to an altered gravity. The other animals were placed in normal gravity after 5 months of hypergravity (HL for Long Hypergravity period). During the first part of the experiment, (weeks 1 to 20) the HL and ROT hamsters were exposedto either centrifugation or rotation. In the second part (week 21 to 34), all groups lived in normal gravity.
Animals were distributed into four groups:
The hamsters of all four groups were 1 month old with a mean weight of 32.5 g for the HS and HL hamsters and 41.8 g for the CON and ROT hamsters at the start of the experiment. They lived in acrylate boxes (22 cm x 37 cm; 3 or 4 animals per box), inside the centrifuge-gondola (HL group), in the center of the centrifuge (ROT group) or in similar housings under normal gravity (CON and HS hamsters). Food and water were available ad libitum and the day-night cycle was reversed (light on 19.00 - 7.00 h) . During a centrifuge stop of 30 min, the hamsters were tested in a laboratory room with dimmed lights.
After testing for 32 weeks, the hamsters were killed for histological and morphological examination of the otoconia. Our findings on these were presented in Sondag et al. (1996b). The body weight was measured twice a week. The experiments were performed in accordance with the Principles of Laboratory Animal Care (NIH publication No. 86-23, revised 1985) and with the recommendations provided in a special licence as required by the Dutch Law on the Use of Animals in Scientific Research. The animal centrifuge consisted of a centrally placed 3.5. kW DC motor drive and 2 horizontally mounted arms (length = 115 cm) with aerated and darkened free-swinging gondolas (length: 110 cm, width: 45 cm, height: 80 cm, length arm + gondola: 194 cm). During centrifugation, the Z-vector was constantly directed towards the floor of the gondola. At a rotation speed of 34,3 rpm a 2.5 G-value was reached at the bottom of the gondola. A video camera was installed inside the gondola to allow animal observation (Sondag et al. 1995). The ROT group was placed in a box located exactly on the rotation axis of the centrifuge. Daily, the centrifuge was stopped for animal care and testing.
Tasks
Activity wheel: To observe locomotor activity of HL hamsters during 2.5 G and of CON and HS hamsters during 1 G, an activity wheel was placed in one of the boxes. Daily, the locomotor activity of one hamster of each group was recorded by means of a bicycle-computer (CC-MT200, Cateye, Osaka, Japan). The locomotor activity of the hamsters was measured by the average speed, running time and the distance covered in one day. The activity of each hamster was assessed once in two weeks. Because of limited space in the center of the centrifuge this test was not assessed for the ROT hamsters.
Gait and stride task: The hamsters were held by their chest and their front and hind paws dipped in a tray containing film developer (Agfa) and had to walk in a small plexiglass walkway (length 43 cm, width 9.5 cm) in which an undeveloped X-ray film (Curix) was laid. The film with the paw prints were dipped in rapid fix (Agfa) for 5 minutes, washed for 10 minutes and dried (method earlier described by de Medinaceli et al. 1982). For evaluation of gait and stride we used the method described by Hruska et al (1979). We measured the distance between consecutive left hind paw prints (stride-length), the distance of the interposed right hindpaw perpendicular to the line connecting the two consecutive left hindpaws (stride-width) and the distance between the ipsilateral fore and hind paw prints (superimposed step). At the same time, the dynamic aspects of the walking behaviour were stored on tape. The videorecorder (Panasonic, NV-FS90) could show intervals of 20 ms (50 frames/s). We counted swing time (elevation and forward movement of the foot) and stance time (placement of the foot upon the floor until the next swing) and calculated the percentage of swing time during one stride (swing time / (stance time + swing time)). This task was assessed once in three weeks.
Rail tasks: The rail (acrylate tubes: length 100 cm, diameter 20 mm), placed approximately 20 cm above ground level, were either fixed to standards (fixed rail task) or were movable connected by elastic cords which were attached to the standards (mobile rail task). Both tubes were covered with tape in order to give the hamster a better grip. A platform was placed at the end of the rail where the hamster could collect sun-flower seeds for ± 5 s. One training day was used to train the hamsters for 5 minutes to walk the full length of the rail. Each hamster had to cross the rail 3 times (trials a, b, c) once a week. After one week of HG, testing on the fixed rail started. After two weeks, testing on the mobile rail started. These tests were assessed once a week.
Swimming in a lane (140 x 10 cm, height of the walls 30 cm, water-depth 25 cm, water temperature approximately 300 C) to test swimming ability and speed. The hamsters had to swim to the end of the lane where they could climb an escape ladder. One extra day was used to train the hamsters for 5 minutes to swim to the ladder. On the testing days, the animals swam 6 trials; 3 in the lane and 3 in the water-maze. The crossing time for the middle part of the lane (length 100 cm) was measured. These tests were assessed once a week.
Orientation ability during swimming in a maze (140 x 70 cm). The shape of the maze was similar to our earlier experiments (Sondag et al. 1995; Sondag et al. 1996a). The hamsters had to find a ladder, located at the opposite end of the maze. The crossing time was measured and the orientation strategy (swimming along the walls or straight through the center of the basin) was observed.
Air-righting reflex and resurfacing: The hamsters were dropped in a supine position from a height of 80 cm into a water basin. To avoid visual input this test was performed under infrared light conditions (870 nm) in which hamsters are unable to use visual information (Vauclair et al. 1977). The hamsters’ behaviour was recorded on video for later analysis. Three succeeding trials were given. We measured the number of correct air rightings responses (landing with feet parallel to the surface of the water) and the time needed to resurface after hitting the water. This task was assessed once in three weeks.
For the crossing times and the falling frequency in the rail tests, the crossing times in the swimming tests, the number of correct air-righting responses and for the time needed to resurface the mean of the 3 trials per animal was calculated. Data of the swimming tests, air-righting and resurfacing were statistically assessed with repeated analysis of variance (ANOVA). The stride, gait and rail tests with repeated analysis of variance with weight as covariates (ANCOVA). Post hoc analysis was done with the Tukey’s honestly significant difference test. We used the statistical software SPSS PC+ 5.0 for our analysis (significance: p<0.05).
Results
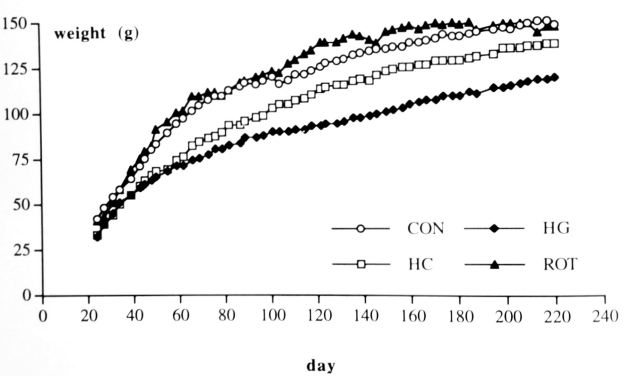
Weight: Repeated measurements showed Group, Time and Group x Time interaction effects in both parts of the experiment. Post hoc analysis revealed that during the whole experiment, the HL and the HS groups weighted less than the CON and the ROT group. Time effects and Group x Time interaction effects showed that all groups increased in weight but the ROT, HS and CON groups increased faster than the HL group (table 1, fig. 1).
Fig. 1. Body weight of the hamsters. HL = hypergravity hamster, HS = HG hamsters after weaning living in 1 G, ROT= rotation hamster and CON = control hamsters. Shown are means.
Treadmill activity: During part 1, Repeated measurements and post hoc analysis revealed that the HL hamsters had a lower average speed, used the treadmill less often and covered less distance than HS and CON hamsters. The results stayed constant for the HL group, while speed, the time of using the treadmill and covered distance decreased in the HS and CON groups (Time effect and Group x Time interaction). In part 2, the average speed and use of treadmill decreased during this period for all three groups (table 1, fig. 2 + 3).
|
part 1 |
part 2 |
|||||
|
Test |
Group |
Time |
Group x Time |
Group |
Time |
Group x Time |
|
Weight |
17.50*** |
1004*** |
12.10*** |
13.83 *** |
53.68 *** |
6.92*** |
|
Treadmill: |
|
|
|
|
|
|
|
Gait: |
|
|
|
|
|
|
|
Fixed rail: |
|
|
|
|
|
|
|
Mobile rail |
|
|
|
|
|
|
|
Swimming: |
|
|
|
|
|
|
|
Air-righting resp. |
110.6*** |
` |
|
|
6.58*** |
|
Table 1: Statistical results of the performance of the hamsters on the perceptive-motor tasks during part 1 and 2. Only significant F- values and p-values are presented; *p<0.05, **p<0.01, ***p<0.001.
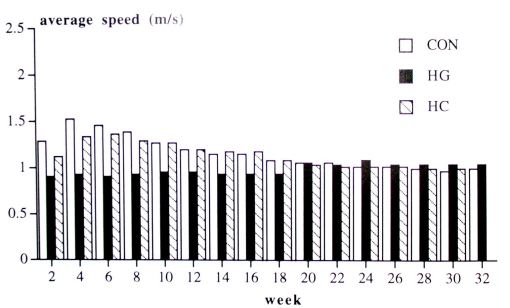
Fig. 2. Treadmill activity: average speed. HL = hypergravity hamster, HS = HG hamsters after weaning living in 1 G, and CON = control hamsters. Shown are means.
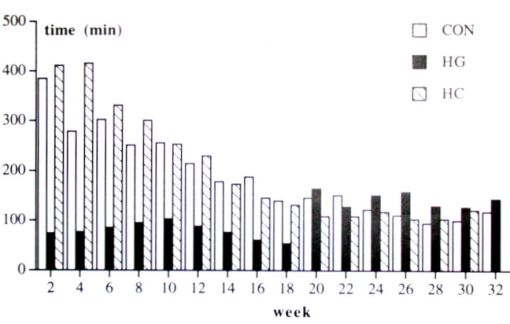
Fig. 3. Treadmill activity: time spent on the treadmill. HL = hypergravity hamster, HS = HG hamsters after weaning living in 1 G, and CON = control hamsters. Shown are means.
Gait and stride test: During the first part of the experiment repeated measurements and post hoc analysis revealed that stride-length and stride-width increased in all groups, faster in the ROT and CON groups than in the HL and HS groups (Time effect and Group x Time interaction, table 1). In the second part, the stride-length decreased for the ROT group and remained constant for the other groups (Group x Time Interaction). Furthermore, repeated measurements revealed that the ROT group showed variability in stance time responses while the other groups showed increases in stance time during this part of the experiment. These stance time responses also affected the percent of swing time (Time effect and Group x Time interaction, table 1).
Fixed rail: 3 of 13 HL hamsters and 2 of 7 ROT hamsters were not able to walk on the rail during the second testing day, whereas all 9 CON hamsters and all 8 HS hamsters could walk on the rail. Concerning crossing time, post hoc testing revealed that during the first 4 weeks the HL hamsters needed more time to cross the rail than the other groups and at the end of the first part the ROT hamsters needed more time than the other groups (Covariate weight almost significant p=0.076). Furthermore, the ROT and HL hamsters fell more from the rail than the CON or HS hamsters and their number of falls increased more than in the other two groups (Group, Time and Group x Time interaction effect, table 1). During the second part, the CON group and ROT needed more time than the HL and HS groups and this time increased more for these groups than for the HL and HS groups at the end of the second part. Furthermore, the ROT group fell more often during the last two months (Group, Time and Group x Time interactions, table 1).
Mobile rail: During the first two months the ROT hamsters needed more time to cross the rail than the other groups and at the end of the first part also the CON hamsters needed more time than the HL and HS groups. Furthermore, the ROT and HL hamsters fell more from the rail than the CON or HS hamsters and their number of falls increased more than in the other two groups. During the second part, the CON group and ROT needed more time than the HL and HS groups. This time remained constant while the crossing time for the HL and HS hamsters increased. All groups increased their falling rate over the last months.
Swimming behaviour in lane and maze: In the lane: the HL hamsters showed disturbances in swimming like circling under water (half of the group had to be saved from drowning), swimming in an almost vertical position and trying to cling on the walls of the lane. At the end of the first part, 7 of 13 HL hamsters (54%) were able to swim to the stair in the lane during the 3 trials. However repeated measurement was impossible for this group because of missing trials during the whole experiment (the hamsters was not able to finish the 3 trials every week). Therefore, their data was excluded in the statistical analysis of the
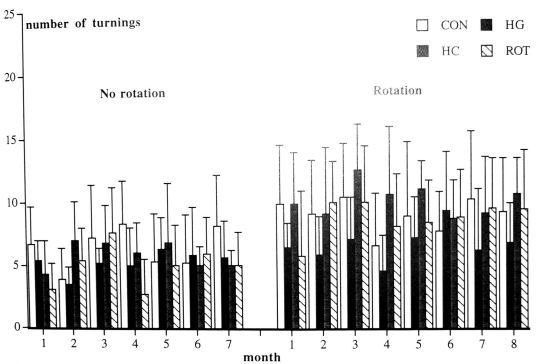
Fig. 4. Number of turnings during a) the no-rotation task, b) the rotation task. HL = hypergravity hamster, HS = HG hamsters after weaning living in 1 G, ROT= rotation hamster and CON = control hamsters. Shown are means and standard deviations.
swimming data. Of the HS hamsters 5 of 8 hamsters (63%) were able to find the stair in the first part. None of the HS hamsters had to be saved from drowning. The CON and ROT hamsters swam straight to the stair. Repeated measurements and posthoc analysis showed that the HS hamsters that were able to find the stair needed more time to swim to the stair than the CON or ROT hamsters (Group effect). The crossing time decreased during this period for all groups (Time effect). In the second part, 5 of 11 HL hamsters (45%) and 5 of 8 HS hamsters (63%) were able to finish three trials in the lane after 3 months of normal gravity. The crossing time of the HS hamsters remained higher than for the CON and ROT groups (Group effect, table 1). In the maze, the 4 of 13 HL hamsters (31%), 5 of 8 HS hamsters (63%), 6 of 7 ROT hamsters (86%) and all 9 CON hamsters were able to find the stair in the first and second part (4 of 11 HL hamsters, 36%) during the 3 succeeding trials. The crossing time decreased for all hamsters during the first part, although faster in the CON and ROT group than in the HS group (Time effect and Group x Time interaction). During the second part, The HS group needed more time to find the stair than the CON and ROT groups. For all three groups the crossing time decreased (Group effect and Group x Time interaction).
Air-rightings reflex: During both parts of the experiment, the HL hamsters made less correct air-righting responses than the other groups (mean number of correct air-rightings: HL hamsters 26.4%, HS hamsters 86%, ROT hamsters 87%, mean CON hamsters 95.3%). At the end of part 2 the number of correct responses increased for the HL hamsters (table 1).
Discussion
To our knowledge this is the first study in which the effects of prenatal development in hypergravity on behaviour were investigated. Other studies dealt with the effects of hypergravity on behaviour in animals which were born in normal gravity after maturation of the sensory systems (Clark, 1974; Fox et al. 1992, Sondag et al. 1996a). We will compare the results obtained from these studies with the results obtained from the present one.
Hamsters conceived and born in hypergravity weighted less than hamsters conceived and born in normal gravity (HL and HS hamsters versus CON and ROT hamsters) which is in accordance with earlier studies concerning the effect of hypergravity on body weight in rodents (Briney and Wunder, 1962; Oyama and Platt, 1967; Serova, 1991). Furthermore, we observed that the HL animals were also smaller in size than CON or ROT hamsters. Rotation in the center of the centrifuge (ROT group) did not result in a body weight decrease, which is in accordance with data coming from other experiments (Martin, 1980). This shows that the increased G-load is responsible for the bodyweight alteration and not the stress caused by continuous rotation during centrifugation. Furthermore, the HL and HS hamsters remained low in body weight and looked smaller in size even after a long period in normal gravity which was also found in hamsters exposed to longterm hypergravity after weaning (Sondag et al. 1996a). This supports the conclusion of Sondag et al. (1996a) that hypergravity leads to a alteration in the basal metabolism. The alteration occurs in both pre-natal and post-natal exposed animals and therefore is independent of embryonal development in normal or hypergravity. Subadult hamsters born in HG (HL and HS group) and hamsters exposed to hypergravity after birth (Sondag et al. 1996a) weighted less than the CON hamsters even after months of normal gravity (up to 8 months for the HS hamsters) suggesting that the basal metabolism after HG exposure is irreversibly altered. Histological research to the cause of these body weight changes showed that especially the bodyfat was decreased in HG adapted animals probably as a result of the higher energy demands placed upon them in hypergravity (Feller et al. 1968).
We suggest that the increased G-load and the additional energy demands were also responsible for the lower treadmill activity (average speed, total running time and distance covered) of the HL group. The hamsters of the HS group showed the same treadmill activity as the CON hamsters. We conclude that hypergravity affects treadmill activity during this period by increasing the energy demands to run the treadmill. When the HL animals lived in normal gravity the differences between the groups disappeared. Therefore, hypergravity does not result in lasting alterations in either the motivation to use the treadmill or in the performance on the treadmill when HG adapted animals are living in normal gravity.
Concerning locomotion in normal gravity, we found that gait and stride were not affected by hypergravity or rotation. The stride-length increased faster for the CON and ROT hamsters than for the the HL and HS hamsters. We suggest that the difference in stride-length can be attributed to the size of the animals. As was noted earlier, animals developed in normal gravity (CON and ROT hamsters), grew faster than the HG developed animals and thus could make larger strides. In the last weeks of the second part (normal gravity) we observed that the ROT hamsters were more often standing still than the hamsters of the other groups causing in an increase in both stance time and percent swing time and a decrease in stride-length during this period. We do not think that this difference can be attributed to a rotation effect.
During the first month of the experiment, balancing on rails was more difficult for the HL and ROT group, causing increases in the crossing times and the number of falls. The ROT hamsters showed a decreased performance on the rail task during the last period of part 1 which can partly be attributed to their higher bodyweight (coviarate body weigth was almost significant). In earlier experiments we found that equilibrium maintenance on small rail was affected by differences in bodyweight resulting in a change of the center of gravity in animals (Sondag et al. 1996a). We conclude that both the HL hamsters and ROT hamsters had difficulties with keeping balance resulting from altered vestibular input caused by either hypergravity or rotational accelerations. In the HL group, these balance problems disappeared because of training experience. However, the ROT hamsters remained to have problems because their faster increasing body size changed their center of gravity, thus making this task even more difficult for them than for the HL hamsters.
Differences in spatial orientation between hamsters living in hypergravity (HL group) and hamsters living in normal gravity (CON and HS groups) were found in the number of correct air-righting responses. The ROT hamsters, subjected to rotatory accelerations, behaved the same as the CON and HS hamsters. We assume that the cause of these disturbances in spatial orientation in the HL animals is the increase in G-load and not the rotatory forces experienced by animals when living in a centrifuge. These disturbances in spatial orientation remained when the HL animals were living in normal gravity (although not significant in the rotation task). The HS hamsters, placed in normal gravity after 21 days, did not show these disturbances. These results show that animals, developed in hypergravity but transferred to normal gravity when they are still young, are able to adapt at least partially to normal gravity.
Persistent disturbances in spatial orientation
were also found in our data concerning the swimming ability of our HL developed
hamsters. We found that our HG developed hamsters (HL and HS group) had severe
disturbances in swimming ability. Rodents postnatally subjected to hypergravity
showed less severe swimming disturbances (Fox et al. 1992; Sondag et al. 1995;
Sondag et al. 1996a) than hamsters conceived and born in hypergravity. The observed
swimming disturbances resembled the swimming deficits also found in rodents
with lesions in the peripheral vestibular system (Petrosini, 1984; Ossenkopp
et al. 1992) and rodents subjected to a decrease of gravity during parabolic
flight (Huygen et al. 1986). We conclude that the swimming deficits we have
found in our HG animals were caused by an alteration in the peripheral vestibular
system. Normal otoconia on the utricle and saccule are necessary for normal
swimming behaviour, and it is suggested that disturbances in swimming behaviour
reflects otoconia defects (Lim and Erway, 1974; Douglas et al. 1979). Earlier,
we reported a structural alteration in the vestibular end organs in these HL
hamsters consisting changes in the relative size of areas with large, medium-sized
or small utricular otoconia and the development of these areas on the utricular
patch (Sondag et al. 1996b). Rotation without the increased G-load did not affect
the swimming ability; the ROT group did not differ from the CON group in swimming
speed and used the same orientation strategies to find the stair in the water-maze
as controls. We did not examine the otoconial layers of these hamsters, but
Krasnov (1991) found no otoconial changes in rats prenatally developed during
continuous rotation (1.1 G) at the center of their centrifuge.
We found that the swimming ability of our HG hamsters did not improve after
a long period of normal gravity. This suggests that alteration in the peripheral
vestibular system was the cause of the swimming deficit. Moreover, the HG animal
is unable to compensate for the ineffective vestibular information by using
either visual or proprioceptive information. Although the swimming ability was
almost as bad as that of the HL hamsters, we found no differences between the
HS group and the CON group on other variables, such as treadmill activity, open
field activity and air-righting responses. The better performance of these HG
developed hamsters during the swimming and air-righting tasks indicate that
there is a critical period for adaptation to a different gravity level. How
this adaptation takes place is unknown. The alterations in the utricular otoconia
distribution of the HL group were also found in the HS group, thus showing no
adaptation to normal gravity (Sondag et al. 1996b). How the behavioural adaptation
to normal gravity takes place in this group remains unclear, but these data
seem to indicate neural plasticity of the young brain which could overcome the
effect of an altered peripheral vestibular system on vestibular evoked behaviour.
We conclude that a structural alteration to hypergravity in the otoconial layers
of the vestibular end organs is one of the causes for the disturbances in air-righting
and swimming found in hamsters prenatally developed and raised in hypergravity.
Our results indicate that these disturbances in spatial orientation become persistent
after a critical period. Furthermore, hypergravity and rotation have a different
effect on vestibular evoked behaviour.
Acknowledgements
The authors gratefully acknowledge the Netherlands Organization for Scientific Research (NWO) for funding this project. This research was conducted while HNPM Sondag was supported by a grant of the Foundation for Behavioural and Educational Sciences (SGW) of this organization (575-62-049), awarded to Prof. Dr. WJ Oosterveld.